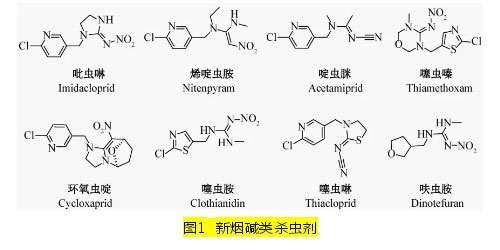
|
| ├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį¼F(xi©żn)ĀŅ╝░Ųõų╬└Ē▓▀┬į |
| üĒį┤Ż║ĪČų▓╬’▒ŻūoīW(xu©”)ł¾ĪĘ2021─ĻĄ┌5Ų┌ 2021-12-14 10:01:00 |
| ├▐č┴Aphis gossypii GloverŻ¼ėųĘQ╣Žč┴Ż¼ī┘░ļ│ß─┐č┴┐ŲŻ¼╩Ūę╗ĘN╩└ĮńąįĘų▓╝Ą─ųžę¬║”ŽxĪŻ├▐č┴─▄ē“×ķ║”├▐╗©Īó─Šķ╚Īó³S╣ŽĪó╠╣ŽĪóŪčūėĪó└▒ĮĘ║═ąĪ¹£Ą╚300ČÓĘNų▓╬’ĪŻū„×ķę╗ĘNųžę¬Ą─┤╠╬³╩Į┐┌Ų„║”ŽxŻ¼├▐č┴┐╔═©▀^ų▒Įė╚Ī╩│ų▓╬’ųŁę║║═é„▓źų▓╬’▓ĪČŠįņ│╔╬Ż║”ĪŻķLŲ┌ęįüĒŻ¼Ę└ų╬├▐č┴ų„ę¬ę└┘ćė┌╗»īW(xu©”)ÜóŽx䮯¼Ą½├▐č┴Š▀ėąĘN╚║öĄ(sh©┤)┴┐┤¾ĪóĘ▒ų│╦┘Č╚┐ņĪó▀mæ¬(y©®ng)ąįÅŖĄ╚╠ž³cŻ¼─▄┐ņ╦┘▀mæ¬(y©®ng)Ę└ų╬╦Ää®▓ó«a(ch©Żn)╔·┐╣╦ÄąįŻ¼ć└(y©ón)ųžė░Ēæ╗»īW(xu©”)Ę└ų╬Ą─ą¦╣¹Ż¼ī”╬ęć°├▐╗©Ė▀┘|(zh©¼)┴┐░▓╚½╔·«a(ch©Żn)įņ│╔ć└(y©ón)ųžĄ─Įø(j©®ng)Ø·ōp╩¦ĪŻą┬¤¤ēAŅÉÜóŽxä®╩ŪĮ³─ĻüĒ├▐č┴╠’ķgĘ└ų╬Ą─ų„ę¬╦Ä䮯¼Ą½├▐č┴ī”įōŅÉ╦Ä䮫a(ch©Żn)╔·▌^Ė▀╦«ŲĮĄ─┐╣ąįŻ¼ęč│╔×ķ├▐č┴ėąą¦Ę└┐žųą├µ┼RĄ─ūŅ┤¾╠¶æ(zh©żn)ĪŻ▒Š╬─Å─├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį¼F(xi©żn)ĀŅĪó┐╣ąįÖCųŲęį╝░┐╣ąįų╬└Ē▓▀┬įĄ╚ĘĮ├µ▀MąąŠC╩÷Ż¼ęįŲ┌×ķ├▐č┴┐╣╦Äąį┐ŲīW(xu©”)ų╬└Ē╠ß╣®ģó┐╝ĪŻ 01Īóą┬¤¤ēAŅÉÜóŽxä® ą┬¤¤ēAŅÉÜóŽxä®╩Ū╚½Ū“ūŅųžę¬Ą─ę╗ŅÉ╗»īW(xu©”)ÜóŽx䮯¼ī”ČÓĘN║”ŽxŠ▀ėą▌^Ė▀Ą─ÜóŽx╗ŅąįŻ¼╩ŪĘ└ų╬č┴ŽxĪó’w╩ŁĪó╚~ŽsĄ╚┤╠╬³╩Į┐┌Ų„║”ŽxūŅėąą¦Ą─ÜóŽxä®ĘNŅÉų«ę╗ĪŻ▒M╣▄įńį┌1690─Ļ╚╦éāŠ═ķ_╩╝ė├¤¤▓▌Į■│÷ę║Ż©ų„ę¬│╔Ęų×ķ¤¤ēAŻ®Ę└ų╬▐r(n©«ng)śI(y©©)║”ŽxŻ¼Ą½ą┬¤¤ēAŅÉÜóŽx䮥─š²╩ĮčąųŲ╩╝ė┌1970─ĻÜż┼Ų╣½╦Šęį¤¤ēAĮY(ji©”)śŗ(g©░u)×ķ╗∙ĄA(ch©│)Ą─Ž╚ī¦(d©Żo)╗»║Ž╬’SD-031588Ą─ķ_░l(f©Ī)ĪŻ1985─ĻŻ¼Ą┬ć°░▌Č·╣½╦Š║═╚š▒Š▐r(n©«ng)╦ÄųĻ╩ĮĢ■╔ń║Žū„ķ_░l(f©Ī)│÷┴╦Ą┌ę╗éĆą┬¤¤ēAŅÉÜóŽxä®Ī¬Ī¬▀┴Žx▀°Ż¼▓óė┌1991─Ļė╔░▌Č·╣½╦Š▀Mąą╚½Ū“═ŲÅVĪŻļS║¾Ż¼ć°ļH╔ŽĄ─Äū╝ę┤¾ą═▐r(n©«ng)╦Ä╣½╦ŠŽÓ└^čą░l(f©Ī)│÷Ž®ÓżŽx░ĘĪóÓżŽxļ▀║═ÓńŽxÓ║Ą╚ČÓĘNą┬¤¤ēAŅÉÜóŽx䮯©łD1Ż®ĪŻ
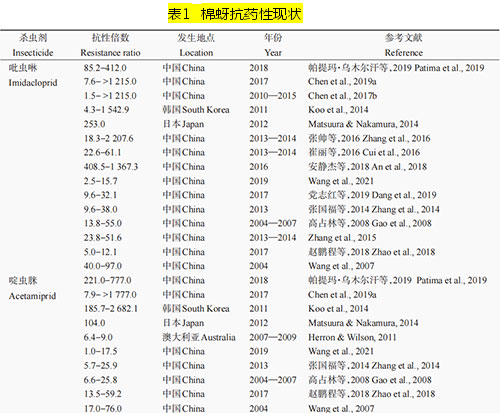
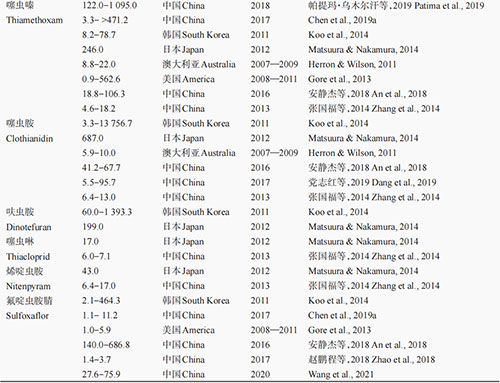
ą┬¤¤ēAŅÉÜóŽxä®ū„ė├ė┌└źŽx¤¤ēAą═ęꧯ─æēA╩▄¾wŻ©nicotinic acetylcholine receptorŻ¼nAChRŻ®Ż¼╩ŪnAChRĄ─▀xō±ąį╝żäė䮯¼įōŅÉÜóŽxä®─▄ē“═©▀^┼c└źŽxnAChRĄ─ĮY(ji©”)║ŽūĶöÓ└źŽxųąśą╔±Įø(j©®ng)ŽĄĮy(t©»ng)ą┼╠¢Ą─š²│Żé„ī¦(d©Żo)Ż¼ī¦(d©Żo)ų┬║”Žx╦└═÷ĪŻą┬¤¤ēAŅÉÜóŽx䮊▀ėąĖ▀ą¦ĪóÅVūVĪóā╚(n©©i)╬³ąį║├Ą─╠ž³cŻ¼╝µŠ▀ė|Üó║═╬ĖČŠū„ė├Ż¼ī”▓Ė╚ķäė╬’Ą═ČŠŻ¼┐╔ėąą¦Ę└ų╬░ļ│ß─┐ĪóŪ╩│ß─┐Īóļp│ß─┐Ą╚ČÓĘN║”ŽxĪŻūį▀┴Žx▀°╔Ž╩ąęįüĒŻ¼ą┬¤¤ēAŅÉÜóŽxä®ę“Ųõ┴╝║├Ą─ÜóŽx╗ŅąįŻ¼į┌╩└ĮńĘČć·ā╚(n©©i)ęč│╔╣”╚Ī┤·┴╦ėąÖC┴ūŅÉĪó░▒╗∙╝ū╦ߧźŅÉĄ╚é„Įy(t©»ng)╦Ää®į┌ū„╬’▒ŻūoųąĄ─æ¬(y©®ng)ė├ĪŻą┬¤¤ēAŅÉÜóŽxä®į┌╚½Ū“╩ął÷ųąš╝ėąųžę¬Ąž╬╗Ż¼į┌2014─ĻįōŅÉÜóŽx䮥─╩ął÷š╝ėą┬╩į┌25%ęį╔ŽŻ¼Į³─ĻüĒŲõį┌╚½Ū“ÜóŽxä®╩ął÷ųąĄ─š╝ėą┬╩ļmėąŽ┬ĮĄŻ¼Ą½╚į╩Ū╚½Ū“Ą┌ę╗┤¾ÜóŽxä®ŅÉą═ĪŻ 02Īó├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį¼F(xi©żn)ĀŅ 2.1 ć°═Ō┐╣╦Äąį¼F(xi©żn)ĀŅ ļSų°ą┬¤¤ēAŅÉÜóŽxä®╩╣ė├┴┐Ą─į÷╝ėŻ¼įńį┌2005─Ļ║”Žxī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįå¢Ņ}Š═ę²Ų┴╦╚╦éāĄ─ĻP(gu©Īn)ūóĪŻ├▐č┴ū„×ķę╗ĘN┐╣ąį▀M╗»║▄┐ņĄ─▐r(n©«ng)śI(y©©)║”ŽxŻ¼ęčī”ČÓĘNą┬¤¤ēAŅÉÜóŽx䮫a(ch©Żn)╔·┴╦┐╣ąįĪŻ▒M╣▄į┌20╩└╝o(j©¼)90─Ļ┤·ą┬¤¤ēAŅÉÜóŽxä®▓┼ķ_╩╝╩ął÷╗»═ŲÅVŻ¼Ą½į┌2000─Ļū¾ėęŠ═ėą╬─½Ił¾Ą└├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮫a(ch©Żn)╔·┴╦┐╣ąįŻ©▒Ē1Ż®ĪŻūį2001─Ļ╩ū┤╬░l(f©Ī)¼F(xi©żn)ęŌ┤¾└¹║═ŲŽ╠čč└├▐č┴ĘN╚║ī”▀┴Žx▀°├¶ĖąąįĮĄĄ═ęįüĒŻ¼├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįå¢Ņ}╚šęµć└(y©ón)ųžĪŻ└²╚ńŻ¼Herron & Wilson═©▀^į\öÓėŗ┴┐Ę©▒O(ji©Īn)£y░l(f©Ī)¼F(xi©żn)Ż¼2006Ī¬2008─Ļ░─┤¾└¹üå├▐č┴╠’ķgĘN╚║ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįų─Ļ╝ėųžŻ¼ų┴2008─Ļęčī”ÓżŽxļ▀ĪóÓńŽx░Ę║═ÓńŽxÓ║Ęųäe«a(ch©Żn)╔·┴╦6.4▒ČĪó10▒Č║═22▒ČĄ─┐╣ąįŻ¼Ūę▀@ĘN┐╣ąį┌ģä▌▀Ćį┌▓╗öÓ░l(f©Ī)š╣Ż¼ĄĮ2009Ī¬2010─Ļ╦∙▒O(ji©Īn)£yĄ─░─┤¾└¹üå├▐č┴ĘN╚║ųą┐╣ąįéĆ¾wĄ─▒╚└²ęčĮø(j©®ng)Ė▀▀_(d©ó)96%ĪŻį┌├└ć°Ż¼2008Ī¬2011─ĻĄ─┐╣╦Äąį▒O(ji©Īn)£yĮY(ji©”)╣¹▒Ē├„Ż¼├▐č┴ī”ÓńŽxÓ║ęčĮø(j©®ng)«a(ch©Żn)╔·┴╦Ė▀╦«ŲĮ┐╣ąįŻ¼Ųõ┐╣ąį▒ČöĄ(sh©┤)ūŅĖ▀ęč▀_(d©ó)562.6▒ČĪŻ2012─ĻŻ¼į┌╚š▒ŠīmŲķ╩ąę▓░l(f©Ī)¼F(xi©żn)▓╔ūį³S╣Ž║═╠ĮĘ╔ŽĄ─├▐č┴ĘN╚║ī”▀┴Žx▀°Īó▀╗Žx░ĘĪóÓńŽx░ĘĪóÓńŽxÓ║ĪóŽ®ÓżŽx░ĘĪóÓżŽxļ▀║═ÓńŽx▀°▀@7ĘNą┬¤¤ēAŅÉÜóŽx䮊∙«a(ch©Żn)╔·┴╦ųąĄ╚ų┴Ė▀Ą╚╦«ŲĮĄ─┐╣ąįŻ¼Ųõųąī”ÓńŽx░ĘĄ─┐╣ąį╦«ŲĮūŅĖ▀×ķ687▒ČŻ¼ī”ÓńŽx▀°┐╣ąį╦«ŲĮūŅĄ═ę▓▀_(d©ó)ĄĮ17▒ČŻ¼Č°ī”ŲõėÓ5ĘNą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį▒ČöĄ(sh©┤)į┌43Ī½253▒Čų«ķgĪŻ═¼śėŻ¼į┌Ēnć°Ż¼2012─ĻĄ─┐╣ąį▒O(ji©Īn)£yĮY(ji©”)╣¹ę▓▒Ē├„Ż¼▓╔ūį└▒ĮĘ║═³S╣Ž╔ŽĄ─├▐č┴ī”6ĘNą┬¤¤ēAŅÉÜóŽx䮊∙«a(ch©Żn)╔·┴╦▌^Ė▀╦«ŲĮĄ─┐╣ąįŻ¼Ųõųąī”ÓżŽxļ▀ĪóÓńŽx░ĘĪó▀┴Žx▀°║═ÓńŽx▀°Ą─┐╣ąį▒ČöĄ(sh©┤)ūŅĖ▀Ęųäe▀_(d©ó)2,682▒ČĪó13,756▒ČĪó1,542▒Č║═1,393▒ČĪŻęį╔ŽčąŠ┐ĮY(ji©”)╣¹▒Ē├„Ż¼├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį╩«Ęųć└(y©ón)ųžŻ¼ęč│╔×ķ╩└ĮńąįļyŅ}ĪŻ
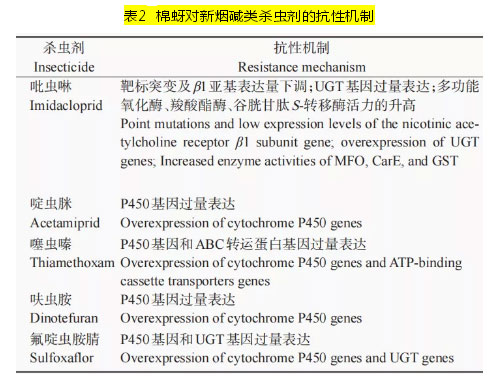
2.2 ć°ā╚(n©©i)┐╣╦Äąį¼F(xi©żn)ĀŅ ć°ā╚(n©©i)├▐č┴ĘN╚║ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįł¾Ą└╔į═Ēė┌ć°═ŌŻ©▒Ē1Ż®ĪŻ2004─ĻŻ¼WangĄ╚▒O(ji©Īn)£y░l(f©Ī)¼F(xi©żn)▓╔ūį╔Į¢|╩ĪĄ┬ų▌╩ąĪó║╩Ø╔╩ąĪó┴─│Ū╩ąĪóĄ┬ų▌╩ąĪó╠®░▓╩ąĄ╚ĄžĄ─├▐č┴╠’ķgĘN╚║ī”▀┴Žx▀°║═ÓżŽxļ▀Ęųäe«a(ch©Żn)╔·┴╦40Ī½97▒Č║═17Ī½76▒ČĄ─┐╣ąįĪŻ2004Ī¬2007─ĻŻ¼║ė▒▒╩Ī└╚Ę╗╩ąĪó▒ŻČ©╩ąĪó£µų▌╩ąĄ╚ĄžĄ─├▐č┴ĘN╚║ī”▀┴Žx▀°Ą─┐╣ąįŠ∙▀_(d©ó)ĄĮ┴╦ųąĄ╚ų┴Ė▀Ą╚╦«ŲĮŻ¼ūŅĖ▀┐╣ąį▒ČöĄ(sh©┤)▀_(d©ó)55▒ČŻ╗└╚Ę╗╩ą║═£µų▌╩ą├▐č┴ĘN╚║ī”ÓżŽxļ▀ę▓«a(ch©Żn)╔·┴╦Ą═Ą╚ų┴ųąĄ╚╦«ŲĮ┐╣ąįŻ¼┐╣ąį▒ČöĄ(sh©┤)ūŅĖ▀▀_(d©ó)25.8▒ČĪŻ2013─ĻŻ¼Åłć°ĖŻĄ╚ī”▓╔ūį╔Į¢|╩Ī┴─│Ū╩ąĪó¢|ĀI╩ą║═╠®░▓╩ąĄ─├▐č┴ĘN╚║ī”6ĘNą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį╦«ŲĮ▀Mąą▒O(ji©Īn)£yŻ¼░l(f©Ī)¼F(xi©żn)╔Į¢|╩Ī├▐č┴ĘN╚║ī”▀┴Žx▀°ĪóÓżŽxļ▀ĪóŽ®ÓżŽx░ĘĪóÓńŽxÓ║║═ÓńŽx░ĘŠ∙«a(ch©Żn)╔·┴╦ųąĄ╚╦«ŲĮ┐╣ąįŻ¼ī”ÓńŽx▀°«a(ch©Żn)╔·Ą═Ą╚╦«ŲĮ┐╣ąįĪŻ2013Ī¬2014─ĻŻ¼Ū±┐hĪó×Ių▌╩ąĪó╬„╚A┐hĄ─├▐č┴ĘN╚║ī”▀┴Žx▀°Ą─┐╣ąį▒ČöĄ(sh©┤)Š∙┤¾ė┌900▒ČŻ¼▀_(d©ó)ĄĮĖ▀╦«ŲĮ┐╣ąįĪŻÅłīW(xu©”)زĄ╚ė┌2010─ĻĄ─▒O(ji©Īn)£yĮY(ji©”)╣¹▒Ē├„Ż¼ą┬Į«ŠS╬ßĀ¢ūÕūįų╬ģ^(q©▒)Ż©║åĘQą┬Į«Ż®╩»║ėūė╩ąĄ─├▐č┴╠’ķgĘN╚║ī”ÓżŽxļ▀«a(ch©Żn)╔·┴╦Ė▀╦«ŲĮ┐╣ąįŻ¼┐╣ąį▒ČöĄ(sh©┤)ūŅĖ▀▀_(d©ó)270.0▒ČĪŻ║ė▒▒╩Ī└╚Ę╗╩ąĪóą┬Į«░ó┐╦╠KĪó╔Į¢|╩ĪĄ┬ų▌╩ą║═ą┬Į«┐³══╩ą├▐č┴ĘN╚║ī”▀┴Žx▀°Ęųäe«a(ch©Żn)╔·┴╦22.6▒ČĪó26.3▒ČĪó53.5▒Č║═61.1▒ČĄ─┐╣ąįĪŻ2018─ĻŻ¼┼┴╠ß¼öĪż×§─ŠĀ¢║╣Ą╚蹊┐░l(f©Ī)¼F(xi©żn)Ż¼▓╔ūįą┬Į«├▐ģ^(q©▒)Ą─├▐č┴ĘN╚║ī”ą┬¤¤ēAŅÉÜóŽx䮫a(ch©Żn)╔·┴╦ųąĪóĖ▀╦«ŲĮĄ─┐╣ąįŻ¼Ųõųąī”▀┴Žx▀°ĪóÓżŽxļ▀║═ÓńŽxÓ║Ą─┐╣ąįĘųäe×ķ85.2Ī½412▒ČĪó221Ī½777▒Č║═122Ī½1,095▒ČĪŻ═¼─ĻŻ¼░▓ņoĮ▄Ą╚▒O(ji©Īn)£yĮY(ji©”)╣¹▒Ē├„Ż¼║ė▒▒╩Ī├▐č┴ĘN╚║ī”▀┴Žx▀°ĪóÓńŽxÓ║ĪóÓńŽx░ĘŠ∙«a(ch©Żn)╔·┴╦ųąĄ╚ų┴Ė▀Ą╚╦«ŲĮ┐╣ąįŻ¼Ųõųąī”▀┴Žx▀°Ą─┐╣ąį╦«ŲĮĖ▀▀_(d©ó)1,367.3▒ČĪŻ2019─ĻŻ¼▓┐Ęų▓╔ūį╔Į¢|╩Ī³S╣Ž╔ŽĄ─╣Žč┴Ż©├▐č┴Ż®ĘN╚║ī”▀┴Žx▀°║═ÓżŽxļ▀«a(ch©Żn)╔·┴╦ųąĄ╚╦«ŲĮ┐╣ąįŻ¼┐╣ąį▒ČöĄ(sh©┤)ūŅĖ▀Ęųäe▀_(d©ó)15.7▒Č║═17.5▒ČĪŻęį╔ŽĮY(ji©”)╣¹▒Ē├„Ż¼╬ęć°├▐č┴╠’ķgĘN╚║ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį╦«ŲĮŲš▒ķ▌^Ė▀Ż¼ąĶ└^└m(x©┤)╝ėÅŖ┐╣ąįų╬└ĒŻ¼ęįčėŠÅ┐╣ąį░l(f©Ī)š╣Ż¼╠ßĖ▀ą┬¤¤ēAŅÉÜóŽxä®ī”├▐č┴Ą─Ę└ų╬ą¦╣¹ĪŻ 2.3 Į╗╗ź┐╣ąį ą┬¤¤ēAŅÉÜóŽxä®ę“Ųõ┼cŲõ╦¹ĘNŅÉÜóŽx䮤oĮ╗╗ź┐╣ąįČ°▒╗ÅVĘ║╩╣ė├Ż¼Ą½įĮüĒįĮČÓĄ─蹊┐░l(f©Ī)¼F(xi©żn)ą┬¤¤ēAŅÉÜóŽxä®ų«ķg┤µį┌▌^Ė▀Ą─Į╗╗ź┐╣ąį’L(f©źng)ļUŻ¼ųĄĄ├┤¾╝ęūóęŌĪŻ└²╚ńŻ¼ChenĄ╚Į╗╗ź┐╣ąį£yČ©ĮY(ji©”)╣¹▒Ē├„Ż¼┐╣▀┴Žx▀°├▐č┴ŲĘŽĄŻ©┐╣ąį▒ČöĄ(sh©┤)ŻŠ1,215▒ČŻ®ī”ÓżŽxļ▀║═Ę·ÓżŽx░ĘļµŠ▀ėąĖ▀▀_(d©ó)1,776▒Č║═260▒ČĄ─Į╗╗ź┐╣ąįĪŻWeiĄ╚░l(f©Ī)¼F(xi©żn)┐╣ÓńŽxÓ║├▐č┴ŲĘŽĄī”┬ō(li©ón)▒ĮŠš§źŻ©11.71▒ČŻ®ĪóĘ·┬╚ŪĶŠš§źŻ©17.90▒ČŻ®ĪóŪĶ╬ņŠš§źŻ©6.85▒ČŻ®ĪóÓńŽx░ĘŻ©6.56▒ČŻ®ĪóÜóōõ┴ūŻ©5.34▒ČŻ®║═Ė▀ą¦┬╚ŪĶŠš§źŻ©4.53▒ČŻ®Š∙┤µį┌├„’@Ą─Į╗╗ź┐╣ąįĪŻZhangĄ╚£yČ©ĮY(ji©”)╣¹ę▓▒Ē├„Ż¼┐╣ÓńŽxÓ║╣Žč┴Ż©├▐č┴Ż®ŲĘŽĄŻ©┐╣ąį▒ČöĄ(sh©┤)ŻŠ2,325.6▒ČŻ®ī”ÓżŽxļ▀Īó▀╗Žx░ĘĪóŽ®ÓżŽx░Ę║═ÓńŽx░ĘĘųäeŠ▀ėą76.6▒ČĪó142.3▒ČĪó299.9▒Č║═311.7▒ČĄ─Į╗╗ź┐╣ąįĪŻ┤╦═ŌŻ¼┐╣▀╗Žx░Ę╣Žč┴Ż©├▐č┴Ż®ŲĘŽĄŻ©┐╣ąį▒ČöĄ(sh©┤)×ķ74.7▒ČŻ®ī”ÓńŽxÓ║Š▀ėą15.3▒ČĄ─Į╗╗ź┐╣ąįĪŻ 03Īó├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįÖCųŲ ├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮫a(ch©Żn)╔·┐╣ąįĄ─ÖCųŲų„ę¬╔µ╝░ĮŌČŠ┤·ųx─▄┴”Ą─į÷ÅŖ║═░ąś╦(bi©Īo)├¶ĖąąįĄ─Ž┬ĮĄŻ©▒Ē2Ż®Ż¼Ž┬├µīóų„ę¬Å─╝Ü(x©¼)░¹╔½╦žP450Īó╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├ĖĄ╚Įķī¦(d©Żo)Ą─┤·ųx┐╣ąį║═░ąś╦(bi©Īo)┐╣ąįā╔ĘĮ├µ▀Mąąšō╩÷ĪŻ 3.1 ┤·ųx┐╣ąį 3.1.1 ╝Ü(x©¼)░¹╔½╦žP450Įķī¦(d©Żo)Ą─┐╣╦Äąį ╝Ü(x©¼)░¹╔½╦žP450╩Ū╔·╬’¾wā╚(n©©i)ūŅųžę¬Ą─ĮŌČŠ┤·ųx├ĖŻ¼į┌║”Žx┐╣╦Äąį«a(ch©Żn)╔·▀^│╠ųąŠ▀ėąĘŪ│Żųžę¬Ą─ū„ė├ĪŻū„×ķČÓ╣”─▄č§╗»├ĖŽĄĄ─ų„ę¬ĮM│╔▓┐ĘųŻ¼╝Ü(x©¼)░¹╔½╦žP450ī”ÜóŽx䮥─┤·ųx─▄┴”ų▒Įėė░Ēæ└źŽxī”ÜóŽx䮥──═╩▄ąįĪŻP450Įķī¦(d©Żo)Ą─ÜóŽxä®┤·ųxĮŌČŠū„ė├╩ŪįSČÓ║”Žxī”ÜóŽx䮫a(ch©Żn)╔·┐╣ąįĄ─ųžę¬ÖCųŲĪŻ─┐Ū░ęčų¬├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį┤¾ČÓ┼cP450Įķī¦(d©Żo)Ą─ĮŌČŠ┤·ųxį÷ÅŖėąĻP(gu©Īn)ĪŻ└²╚ńŻ¼╩»ŠwĖ∙═©▀^į÷ą¦ä®įć“×║═├Ė╗Ņąį£yČ©įć“ׯ¼░l(f©Ī)¼F(xi©żn)ČÓ╣”─▄č§╗»├Ėį┌├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįųąŲų°ųžę¬ū„ė├ĪŻ╣∙╠ņ°PĄ╚═¼śė░l(f©Ī)¼F(xi©żn)ČÓ╣”─▄č§╗»├Ė┼c├▐č┴ī”▀┴Žx▀°║═ÓżŽxļ▀Ą─┐╣ąį├▄ŪąŽÓĻP(gu©Īn)ĪŻWuĄ╚═©▀^ī”P450╗∙ę“▒Ē▀_(d©ó)┴┐Ą─Ęų╬÷░l(f©Ī)¼F(xi©żn)Ż¼CYP3╝ęūÕP450╗∙ę“Ą─╔Žš{(di©żo)▒Ē▀_(d©ó)┐╔─▄╩Ū├▐č┴ī”ÓńŽxÓ║«a(ch©Żn)╔·┐╣ąįĄ─ų„ę¬įŁę“ĪŻ═¼śėŻ¼CYP6CY14ĪóCYP6DC1║═CYP6CZ1Ą╚P450╗∙ę“Ą─▀^▒Ē▀_(d©ó)╩Ū├▐č┴ī”ÓżŽxļ▀«a(ch©Żn)╔·┤·ųx┐╣ąįĄ─ųžę¬ÖCųŲŻ¼CYP6CY14ĪóCYP6CY22║═CYP6UN1Ą─▀^▒Ē▀_(d©ó)į┌├▐č┴ī”▀╗Žx░Ę«a(ch©Żn)╔·┐╣ąį▀^│╠ųąŲųžę¬ū„ė├ĪŻ┤╦═ŌŻ¼HirataĄ╚═©▀^S2╝Ü(x©¼)░¹šµ║╦▒Ē▀_(d©ó)į┌ą┬¤¤ēAŅÉÜóŽxä®┐╣ąįŲĘŽĄųą’@ų°╔Žš{(di©żo)▒Ē▀_(d©ó)Ą─CYP6CY22║═CYP6CY13╗∙ę“Ż¼▓ó▀Mąą¾w═Ō┤·ųxįć“ׯ¼░l(f©Ī)¼F(xi©żn)├▐č┴P450─▄ē“┐ņ╦┘┤·ųx▀┴Žx▀°ĪóÓżŽxļ▀ĪóÓńŽx▀°Ą╚ą┬¤¤ēAŅÉÜóŽx䮯¼ūCīŹ┴╦P450╗∙ę“▀^▒Ē▀_(d©ó)╩ŪĮķī¦(d©Żo)├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮫a(ch©Żn)╔·┤·ųx┐╣ąįĄ─ųžę¬ÖCųŲĪŻ 3.1.2 ¶╚╦ߧź├ĖĮķī¦(d©Żo)Ą─┐╣╦Äąį ¶╚╦ߧź├Ėū„×ķ┴Ē═Ōę╗ĘN╩«Ęųųžę¬Ą─ĮŌČŠ├ĖŻ¼─▄ē“┤▀╗»╦«ĮŌ║¼¶╚╗∙§źµIĪó§Ż░ʵI║═┴“§źµIĄ─╗»║Ž╬’ĪŻįSČÓė├ė┌├▐č┴Ę└ų╬Ą─╗»īW(xu©”)ÜóŽx䮯¼╚ńėąÖC┴ūŅÉĪó░▒╗∙╝ū╦ߧźŅÉĪóöM│²ŽxŠš§źŅÉĄ╚ÜóŽx䮯¼ŲõĘųūėĮY(ji©”)śŗ(g©░u)ųąČ╝║¼ėą§źµIŻ¼ę“┤╦Ż¼¶╚╦ߧź├Ė┤·ųx─▄┴”Ą─į÷ÅŖę▓╩Ū├▐č┴┐╣╦Äąį«a(ch©Żn)╔·Ą─ųžę¬ÖCųŲų«ę╗ĪŻęčėąčąŠ┐▒Ē├„Ż¼¶╚╦ߧź├Ė╗ŅąįŻ©ęį”┴NA×ķĄū╬’Ż®į┌├▐č┴▀┴Žx▀°┐╣ąįŲĘŽĄųąŠ∙’@ų°Ė▀ė┌├¶ĖąŲĘŽĄŻ¼═Ų£yŲõ┼c├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįėąĻP(gu©Īn)ĪŻŚŅ¤©ŪÓĄ╚蹊┐░l(f©Ī)¼F(xi©żn)Ż¼ī”▀┴Žx▀°«a(ch©Żn)╔·24.38▒Č┐╣ąįĄ─├▐č┴ŲĘŽĄĄ─¶╚╦ߧź├Ė▒╚╗Ņ┴”╩Ū├¶ĖąŲĘŽĄĄ─2.73▒ČŻ¼╩ĘĢį▒¾Ą╚ę▓░l(f©Ī)¼F(xi©żn)▀┴Žx▀°┐╣ąį├▐č┴ŲĘŽĄĄ─¶╚╦ߧź├Ė▒╚╗Ņ┴”╩Ū├¶ĖąŲĘŽĄĄ─2.87▒ČŻ¼šf├„├▐č┴ī”▀┴Žx▀°Ą─┐╣ąį«a(ch©Żn)╔·┼c¶╚╦ߧź├ĖĮŌČŠ─▄┴”Ą─╔²Ė▀ėąĻP(gu©Īn)ĪŻ┤▐¹ÉĄ╚ī”╠’ķg├▐č┴┐╣ąįŲĘŽĄĄ─├Ė╗Ņąį£yČ©ĮY(ji©”)╣¹▒Ē├„Ż¼ī”▀┴Žx▀°«a(ch©Żn)╔·ųąĄ╚╦«ŲĮ┐╣ąįĄ─├▐č┴╠’ķgĘN╚║Ż©░ó┐╦╠KĘN╚║║═┐³══ĘN╚║Ż®¶╚╦ߧź├ĖĄ─▒╚╗Ņ┴”Ęųäe╩Ū├¶ĖąĘN╚║Ą─1.6▒Č║═1.7▒ČŻ¼šf├„¶╚╦ߧź├Ė╗Ņ┴”Ą─╔²Ė▀┼c├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįą╬│╔ėąĻP(gu©Īn)ĪŻųĄĄ├ūóęŌĄ─╩ŪŻ¼ą┬¤¤ēAŅÉ╦Ä䮥─╗»īW(xu©”)ĮY(ji©”)śŗ(g©░u)ųą▓╗║¼§źµIŻ¼─Ū├┤¶╚╦ߧź├Ė╚ń╣¹ū„×ķ▀┴Žx▀°Ą─┐╣ąįÖCųŲĄ─įÆāHėąę╗ĘN┐╔─▄ąįŻ¼╝┤ū„×ķĮY(ji©”)║ŽĄ░░ūŲū„ė├ĪŻĄ½╩Ū╔Ž╩÷蹊┐ł¾Ą└āH╩Ūę╗éĆŽÓĻP(gu©Īn)ąįŻ¼▓óø]ūC├„ū„×ķĮY(ji©”)║ŽĄ░░ūĄ─ū„ė├ĪŻę“┤╦Ż¼¶╚╦ߧź├ĖĮķī¦(d©Żo)├▐č┴ī”ą┬¤¤ēAŅÉÜóŽxä®┐╣ąįĄ─ĘųūėÖCųŲ▓ó▓╗├„┤_ĪŻ 3.1.3 ╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├ĖĮķī¦(d©Żo)Ą─┐╣╦Äąį ╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├Ė┤▀╗»╣╚ļūĖ╩ļ─ĮY(ji©”)║ŽĘ┤æ¬(y©®ng)ūŅķ_╩╝Ą─▓Į¾EŻ¼╩ŪįōĘ┤æ¬(y©®ng)Ą─ĻP(gu©Īn)µI├ĖŻ¼┤▀╗»ėH║╦ąįŻ©▀ĆįŁąįŻ®Ą─╣╚ļūĖ╩ļ─┼c▓╗═¼ĘNŅÉĄ─ėHļŖūė═Ōį┤╗»║Ž╬’░l(f©Ī)╔·Ę┤æ¬(y©®ng)Ż¼╩╣▀ĆįŁąį╣╚ļūĖ╩ļ─┼cėą║”ėHļŖūė╗∙łFĮY(ji©”)║Ž▓ó┼┼│÷¾w═ŌŻ¼ŲĄĮĮŌČŠ┤·ųxĄ─ū„ė├ĪŻęį╣╚ļūĖ╩ļ─×ķĮķ┘|(zh©¼)Ą─┤·ųxŻ¼ų„ę¬ėą2ĘNŻ║Ą┌1ĘN╩Ūęį▀ĆįŁą═╣╚ļūĖ╩ļ─ū„×ķ┤▀╗»ä®Ą─┤·ųxĘ┤æ¬(y©®ng)Ż¼Ą┌2ĘN╩Ū▀ĆįŁą═╣╚ļūĖ╩ļ─┼cĄū╬’ų▒ĮėĮY(ji©”)║ŽĄ─┤·ųxĘ┤æ¬(y©®ng)ĪŻėąčąŠ┐▒Ē├„╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├Ė┼c├▐č┴ī”ą┬¤¤ēAŅÉÜóŽxä®┐╣ąįŽÓĻP(gu©Īn)Ż¼└²╚ńŻ¼┼╦╬─┴┴Ą╚═©▀^├Ė╗Ņąį£yČ©įć“×ūCīŹ╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├Ė╗Ņ┴”į÷ÅŖį┌├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįųąŲųžę¬ū„ė├ĪŻŚŅ¤©ŪÓĄ╚Ż©2009Ż®čąŠ┐ę▓░l(f©Ī)¼F(xi©żn)Ż¼┐╣▀┴Žx▀°├▐č┴ŲĘŽĄ╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├ĖĄ─▒╚╗Ņ┴”╩Ū├¶ĖąŲĘŽĄĄ─1.57▒ČŻ¼šf├„╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├Ė▒╚╗Ņ┴”Ą─╠ßĖ▀╩Ūę²Ų├▐č┴ī”▀┴Žx▀°«a(ch©Żn)╔·┐╣ąįĄ─ųžę¬įŁę“ų«ę╗ĪŻ┤▐¹ÉĄ╚šJ(r©©n)×ķŻ¼╬ęć°ą┬Į«░ó┐╦╠KĄžģ^(q©▒)║═┐³══╩ąĄ─├▐č┴╠’ķgĘN╚║ī”▀┴Žx▀°Ą─┐╣ąį┼c╣╚ļūĖ╩ļ─S-▐D(zhu©Żn)ęŲ├Ė╗ŅąįĄ─į÷ÅŖŽÓĻP(gu©Īn)ĪŻ 3.1.4 UDP-ŲŽ╠Ū╗∙▐D(zhu©Żn)ęŲ├ĖĮķī¦(d©Żo)Ą─┐╣╦Äąį UDP-ŲŽ╠Ū╗∙▐D(zhu©Żn)ęŲ├ĖŻ©UDP-glycosyltransferaseŻ¼UGTŻ®╩Ūę╗ĘNųžę¬Ą─ó“ŽÓ┤·ųx├ĖŻ¼─▄ē“┤▀╗»ČÓĘNā╚(n©©i)į┤║══Ōį┤╬’┘|(zh©¼)┼cŲŽ╠č╠Ū╚®╦ßĮY(ji©”)║ŽŻ¼į┌└źŽxā╚(n©©i)į┤║══Ōį┤╗»║Ž╬’Ą─ĮŌČŠ┤·ųxųąŠ▀ėąųžę¬ū„ė├ĪŻĮ³─ĻüĒŻ¼ėąčąŠ┐▒Ē├„UGTį┌║”ŽxųąŲĘŪ│Żųžę¬Ą─ĮŌČŠ┤·ųxū„ė├Ż¼ģó┼c║”Žxī”Ę└ų╬╦Ä䮥─┐╣ąįą╬│╔ĪŻ└²╚ńŻ¼LiĄ╚蹊┐░l(f©Ī)¼F(xi©żn)UGTį┌ąĪ▓╦ČĻP(gu©Īn)lutella xylostellaŻ©L.Ż®ī”┬╚Žx▒Į╝ū§Ż░Ę┐╣ąįą╬│╔ųąŲų°ųžę¬ū„ė├ĪŻį┌├▐č┴ųąŻ¼UGTę▓▒╗░l(f©Ī)¼F(xi©żn)į┌├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį«a(ch©Żn)╔·▀^│╠ųąŲų°ųžę¬Ą─ĮŌČŠ┤·ųxū„ė├ĪŻ└²╚ńŻ¼į┌├▐č┴ī”ÓńŽxÓ║┐╣ąįŲĘŽĄųąėą13éĆUGT╗∙ę“’@ų°╔Žš{(di©żo)▒Ē▀_(d©ó)Ż¼ŪęRNAi│┴─¼UGT348A2ĪóUGT344B4║═UGT344J2▀@3éĆUGT╗∙ę“▒Ē▀_(d©ó)Š∙─▄’@ų°╠ßĖ▀┐╣ąį├▐č┴ī”ÓńŽxÓ║Ą─├¶ĖąąįŻ¼šf├„UGT╗∙ę“Ą─▀^┴┐▒Ē▀_(d©ó)┼c├▐č┴ī”ÓńŽxÓ║Ą─┐╣ąįŽÓĻP(gu©Īn)ĪŻChenĄ╚蹊┐░l(f©Ī)¼F(xi©żn)Ż¼į┌├▐č┴▀┴Žx▀°┐╣ąįŲĘŽĄųąUGT343A4ĪóUGT344B4ĪóUGT344C7Ą╚ČÓéĆUGT╗∙ę“’@ų°╔Žš{(di©żo)▒Ē▀_(d©ó)Ż¼Ūęį┌Ą░░ū╦«ŲĮ╔ŽUGT344A16ĪóUGT344B4ĪóUGT344C7║═UGT344N4Ą─▒Ē▀_(d©ó)╦«ŲĮę▓’@ų°Ė▀ė┌├¶ĖąŲĘŽĄŻ¼▀Mę╗▓ĮRNAi│┴─¼UGT344B4║═UGT344C7Ą─▒Ē▀_(d©ó)─▄ē“’@ų°╠ßĖ▀▀┴Žx▀°┐╣ąįŲĘŽĄ├▐č┴ī”▀┴Žx▀°Ą─├¶ĖąąįŻ¼ūCīŹUGT344B4║═UGT344C7Ą─▀^┴┐▒Ē▀_(d©ó)┼c├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįėąĻP(gu©Īn)ĪŻMaĄ╚į┌├▐č┴ī”Ę·ÓżŽx░Ęļµ┐╣ąįÖCųŲ蹊┐ųąę▓░l(f©Ī)¼F(xi©żn)Ż¼UGT344B4ĪóUGT344C5ĪóUGT344A11Ą╚7éĆUGT╗∙ę“į┌├▐č┴Ę·ÓżŽx░Ęļµ┐╣ąįŲĘŽĄųą’@ų°╔Žš{(di©żo)▒Ē▀_(d©ó)Ż¼Č°RNAi│┴─¼4éĆUGT344üå╝ęūÕ╗∙ę“Ż©UGT344B4ĪóUGT344C5ĪóUGT344A11║═UGT344L2Ż®Š∙─▄ē“’@ų°╠ßĖ▀├▐č┴ī”Ę·ÓżŽx░ĘļµĄ─├¶ĖąąįŻ¼šf├„ČÓéĆUGT╗∙ę“Ą─▀^▒Ē▀_(d©ó)ę▓╩Ū├▐č┴ī”Ę·ÓżŽx░Ęļµ«a(ch©Żn)╔·┤·ųx┐╣ąįĄ─ųžę¬įŁę“ų«ę╗ĪŻ 3.1.5 ABC▐D(zhu©Żn)▀\Ą░░ūĮķī¦(d©Żo)Ą─┐╣╦Äąį PanĄ╚蹊┐░l(f©Ī)¼F(xi©żn)Ż¼┼c├¶ĖąŲĘŽĄŽÓ▒╚Ż¼├▐č┴ī”ÓńŽxÓ║┐╣ąįŲĘŽĄųąČÓéĆABC▐D(zhu©Żn)▀\Ą░░ū╗∙ę“’@ų°╔Žš{(di©żo)▒Ē▀_(d©ó)Ż¼ŪęęųųŲ▀^▒Ē▀_(d©ó)Ą─ABC▐D(zhu©Żn)▀\Ą░░ū╗∙ę“ABCA2ĪóABCD1ĪóABCD2ĪóABCE1║═ABCG15─▄ē“’@ų°╠ßĖ▀┐╣ąį├▐č┴ī”ÓńŽxÓ║Ą─├¶ĖąąįŻ¼šf├„├▐č┴ī”ÓńŽxÓ║Ą─┐╣ąį┼cABC▐D(zhu©Żn)▀\Ą░░ūėąĻP(gu©Īn)ĪŻŅÉ╦ŲĄžŻ¼WangĄ╚═©▀^▒╚▌^╩ęā╚(n©©i)├¶ĖąŲĘŽĄ║═Ę·ÓżŽx░Ęļµ┐╣ąįŲĘŽĄA(ch©│)BC▐D(zhu©Żn)▀\Ą░░ū╗∙ę“Ą─▒Ē▀_(d©ó)▓Ņ«ÉŻ¼░l(f©Ī)¼F(xi©żn)AB®\CA1ĪóABCA2ĪóABCB1Ą╚10éĆABC▐D(zhu©Żn)▀\Ą░░ū╗∙ę“į┌┐╣ąįŲĘŽĄųą’@ų°╔Žš{(di©żo)▒Ē▀_(d©ó)Ż¼ŪęRNAĖ╔ö_ABCA1║═ABCD1ā╔éĆ╗∙ę“’@ų°╠ßĖ▀┴╦┐╣ąį├▐č┴ī”Ę·ÓżŽx░ĘļµĄ─├¶ĖąąįŻ¼šf├„ABC▐D(zhu©Żn)▀\Ą░░ū╗∙ę“Ą─╔Žš{(di©żo)▒Ē▀_(d©ó)ę▓ģó┼c┴╦├▐č┴ī”Ę·ÓżŽx░ĘļµĄ─┐╣ąįą╬│╔ĪŻ 3.2 ░ąś╦(bi©Īo)┐╣ąį Ęųūė░ąś╦(bi©Īo)┐╣ąįūā«É╩Ū║”Žx«a(ch©Żn)╔·┐╣╦ÄąįĄ─ųžę¬ÖCųŲĪŻą┬¤¤ēAŅÉÜóŽx䮥─ū„ė├░ąś╦(bi©Īo)╩Ū¤¤ēAą═ęꧯ─æēA╩▄¾wŻ¼ę“┤╦Ż¼nAChR░ąś╦(bi©Īo)╬╗³c═╗ūā╩Ū├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮫a(ch©Żn)╔·┐╣ąįĄ─ųžę¬ÖCųŲĪŻKooĄ╚蹊┐░l(f©Ī)¼F(xi©żn)├▐č┴¤¤ēAą═ęꧯ─æēA╩▄¾w”┬1üå╗∙Łh(hu©ón)Dģ^(q©▒)Ą─R81T═╗ūā╩ŪŲõī”▀┴Žx▀°«a(ch©Żn)╔·┐╣ąįĄ─ųžę¬ÖCųŲŻ¼Ūę¾w═Ōįć“×ūCīŹR81T═╗ūāĮķī¦(d©Żo)Ą─░ąś╦(bi©Īo)├¶ĖąąįŽ┬ĮĄ┼c├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįų▒ĮėŽÓĻP(gu©Īn)ĪŻKimĄ╚į┌├▐č┴▀┴Žx▀°┐╣ąįŲĘŽĄųąÖz£yĄĮL80S║═R81T═╗ūāŻ¼ę▓ūCīŹŲõ┼c▀┴Žx▀°┐╣ąįŽÓĻP(gu©Īn)ĪŻZhangĄ╚▓╔ė├īŹĢr¤╔╣ŌČ©┴┐╠ž«ÉąįĄ╚╬╗╗∙ę“PCRŻ©real-time PCR amplififi cation of specifific-alleleŻ¼RT-PASAŻ®╝╝ąg(sh©┤)Öz£y░l(f©Ī)¼F(xi©żn)Ż¼▓╔ūį└╚Ę╗╩ą║═Ą┬ų▌╩ąĄ─╠’ķg▀┴Žx▀°┐╣ąį├▐č┴ĘN╚║ųą┤µį┌R81T═╗ūāéĆ¾wŻ¼Č°┤▐¹ÉĄ╚═©▀^ī”╬ęć°╠’ķg├▐č┴ĘN╚║Ą─═╗ūāÖz£yę▓░l(f©Ī)¼F(xi©żn)Ż¼nAChR ”┬1üå╗∙Ą─R81T═╗ūā┼c├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįŽÓĻP(gu©Īn)ĪŻChenĄ╚į┌╩ęā╚(n©©i)║Y▀xĄ─▀┴Žx▀°Ė▀┐╣ŲĘŽĄŻ©┐╣ąį▒ČöĄ(sh©┤)ŻŠ1,215▒ČŻ®Ą─”┬1üå╗∙╔Ž═¼ĢrÖz£yĄĮ┴╦R81TĪóV62I║═K264E╚²éĆ╬╗³c═╗ūāŻ¼ŲõųąV62I║═K264E═╗ūā?y©Łu)ķ╩ū┤╬░l(f©Ī)¼F(xi©żn)Ż¼V62I╬╗ė┌LoopDŪ░ęĒą“┴ąŻ¼Č°K264E╬╗ė┌┐ń─żģ^(q©▒)ė“TM1║═TM2ų«ķgŻ¼▀Mę╗▓Įī”╠’ķgĘN╚║Ą─Öz£yĮY(ji©”)╣¹ę▓ūCīŹ▀@3éĆ╬╗³c═╗ūāŠ∙┼c├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįŽÓĻP(gu©Īn)Ż¼šf├„nAChR░ąś╦(bi©Īo)═╗ūā╩Ū├▐č┴ī”▀┴Žx▀°«a(ch©Żn)╔·┐╣ąįĄ─ĻP(gu©Īn)µIįŁę“ĪŻ┤╦═ŌŻ¼MunkhbayarĄ╚║═WangĄ╚═©▀^ī”▓╔ūį╬ęć°ą┬Į«║═╔Į¢|╩ĪĄ─├▐č┴╠’ķgĘN╚║Ą─═╗ūāÖz£yę▓░l(f©Ī)¼F(xi©żn)nAChR”┬1üå╗∙╔ŽR81T║═K264E═╗ūā┼c├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįėąĻP(gu©Īn)ĪŻ ┤╦═ŌŻ¼nAChR╩▄¾w╗∙ę“Ą═╦«ŲĮ▒Ē▀_(d©ó)┐╔─▄ę▓į┌├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįą╬│╔ųą░l(f©Ī)ō]ų°ųžę¬ū„ė├ĪŻ└²╚ńŻ¼▒╚▌^▀┴Žx▀°┐╣ąį║═├¶ĖąŲĘŽĄnAChR”┬1üå╗∙Ą─▒Ē▀_(d©ó)┴┐Ż¼░l(f©Ī)¼F(xi©żn)Ųõ▒Ē▀_(d©ó)┴┐į┌├▐č┴▀┴Žx▀°┐╣ąįŲĘŽĄųą’@ų°Ž┬š{(di©żo)Ż¼šf├„ŲõĄ═╦«ŲĮ▒Ē▀_(d©ó)┼c├▐č┴ī”▀┴Žx▀°Ą─┐╣ąįą╬│╔ėąĻP(gu©Īn)ĪŻ═¼śėŻ¼į┌├▐č┴ÓńŽxÓ║┐╣ąįŲĘŽĄųąŻ¼į┌nAChRĖ„üå╗∙ųąŠ∙╬┤Öz£yĄĮ░ąś╦(bi©Īo)╬╗³c═╗ūāŻ¼Ą½Ųõ”┴1Īó”┴4-1Īó”┴4-2Īó”┴5╝░”┴7üå╗∙į┌▐D(zhu©Żn)õø╦«ŲĮ╔ŽĘųäeŽ┬š{(di©żo)▒Ē▀_(d©ó)3.32▒ČĪó1.60▒ČĪó2.05▒ČĪó5.41▒Č║═1.48▒ČŻ¼šf├„├▐č┴”┴1Īó”┴4-1Īó”┴4-2Īó”┴5╝░”┴7üå╗∙Ą─Ž┬š{(di©żo)▒Ē▀_(d©ó)┐╔─▄į┌├▐č┴ī”ÓńŽxÓ║Ą─┐╣ąįą╬│╔ųąŲų°ę╗Č©ū„ė├ĪŻ
04Īó├▐č┴┐╣ąįų╬└Ē▓▀┬į Georghiou & SaitoÅ─╗»īW(xu©”)╦Ää®╩╣ė├Ą─ĮŪČ╚╠ß│÷┴╦║”Žx┐╣╦Äąįų╬└ĒĄ─3ĘN▓▀┬įŻ¼╝┤▀mČ╚ų╬└ĒĪó’¢║═ų╬└Ē║═ČÓŽ“▀M╣źų╬└ĒĪŻ▀mČ╚ų╬└Ēų„ę¬═©▀^ĮĄĄ═ÜóŽx╦Ä䮥─▀xō±ē║┴”Ż¼čėŠÅ┐╣ąį░l(f©Ī)š╣ĪŻŠ▀¾w┤ļ╩®░³└©▓╔ė├Ą═ä®┴┐ė├╦ÄŻ¼▒Ż┴¶ę╗Č©▒╚└²Ą─├¶Ėą╗∙ę“ą═Ż¼£p╔┘╩®╦Ä┤╬öĄ(sh©┤)Ż¼▀xė├Üłą¦Ų┌Č╠Ą─╗»║Ž╬’Ż¼▒▄├Ō╩╣ė├ŠÅßī䮯¼ų„ę¬ßśī”│╔Žx╩®╦ÄŻ¼▒M┴┐Šų▓┐Č°▓╗╩Ū┤¾├µĘe╩®╦ÄŻ¼╚╦×ķįO(sh©©)ų├ę╗Č©Ą─▒ėūoģ^(q©▒)Ż¼╠ßĖ▀╩®╦Ä║”ŽxĘN╚║ķōųĄĄ╚ĪŻ║═▀mČ╚ų╬└ĒŽÓĘ┤Ż¼’¢║═ų╬└ĒĄ──┐Ą─╩Ū▒M┐╔─▄Ą─╠ßĖ▀▀xō±ē║Ż¼Ž¹│²┐╣ąįļs║ŽūėųąĄ─┐╣ąįĄ╚╬╗╗∙ę“Ż¼čėŠÅ┐╣ąį░l(f©Ī)š╣ĪŻŲõų„ę¬┤ļ╩®╩Ū═©▀^Ė▀ä®┴┐ė├╦ÄŻ¼Üó╦└┤¾ČÓöĄ(sh©┤)ļs║ŽūėŻ¼╩╣┐╣ąį╣”─▄ļ[ąį╗»ĪŻČÓŽ“▀M╣źų╬└Ēų„ę¬╩ŪĖ∙ō■(j©┤)ČŠ╬’ī”╔·╬’Ą─ČÓ╬╗³cū„ė├Ż¼╩╣░ąś╦(bi©Īo)▓╗ęū«a(ch©Żn)╔·┐╣ąįŻ¼ę╗░Ń═©▀^ÜóŽx╦Ä䮥─▌åė├Īó╗ņė├ęį╝░Ęųģ^(q©▒)╩®╦ÄüĒčėŠÅ┐╣ąį░l(f©Ī)š╣ĪŻßśī”├▐č┴┐╣ąįć└(y©ón)ųžĄ─╠ž³cŻ¼Į©ūh▓╔╚ĪęįŽ┬ÄūĘN┤ļ╩®Ż¼▒M┴┐čėŠÅ├▐č┴Ą─┐╣ąį░l(f©Ī)š╣ĪŻ 4.1 ╝ėÅŖ╠’ķgĘN╚║┐╣╦ÄąįäėæB(t©żi)▒O(ji©Īn)£y ┐╣╦Äąį▒O(ji©Īn)£y╩ŪųŲČ©║”Žx┐╣╦Äąįų╬└Ē▓▀┬į║═Š½£╩(zh©│n)▀x╦ÄĄ─ĻP(gu©Īn)µIŻ¼ę¬ŠC║Ž└¹ė├Ė„ĘN┐╣╦ÄąįÖz£y╝╝ąg(sh©┤)Ż¼ŽĄĮy(t©»ng)£yČ©├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį░l(f©Ī)╔·Ņl┬╩║═┐╣ąį╦«ŲĮĄ─Ģr┐šūā╗»Ż¼┴╦ĮŌŲõ┐╣ąį░l(f©Ī)╔·║═░l(f©Ī)š╣Ą─ęÄ(gu©®)┬╔ĪŻ×ķ▒M┴┐čėķLą┬ÜóŽx䮥─╩╣ė├ē█├³Ż¼æ¬(y©®ng)įōÅ─Ųõķ_╩╝æ¬(y©®ng)ė├ų«ĢrŠ═ųŲČ©┐╣ąį▒O(ji©Īn)£yėŗäØŻ¼▓ó│ų└m(x©┤)▀Mąą▒O(ji©Īn)£yĪŻ═¼ĢrŻ¼└¹ė├¼F(xi©żn)┤·Ęųūė╔·╬’īW(xu©”)╝╝ąg(sh©┤)Ż¼š{(di©żo)▓ķ├▐č┴ī”ą┬¤¤ēAŅÉÜóŽxä®┐╣ąįą╬│╔Ą─ŽÓĻP(gu©Īn)╗∙ę“┘Yį┤Ż¼┴╦ĮŌ┐╣ąįŽÓĻP(gu©Īn)╗∙ę“Ą─ĘNŅÉĪóŅl┬╩╝░ŲõĘų▓╝ĪŻĖ∙ō■(j©┤)┐╣ąį▒O(ji©Īn)£yĮY(ji©”)╣¹Ż¼ųŲČ©┐╣ąįų╬└Ē▓▀┬įŻ¼ųĖī¦(d©Żo)╠’ķg┐ŲīW(xu©”)ė├╦ÄŻ¼čėŠÅ├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį░l(f©Ī)š╣Ż¼čėķLÜóŽx䮥─╩╣ė├ē█├³ĪŻ 4.2 ║Ž└Ē▀xō±╗ņė├║═▌åė├ÜóŽxä®ĮM║Ž ┐╣ąįų╬└ĒųąĄ─╦Ää®╗ņė├╩Ū└¹ė├ę╗ĮMŠ▀ėą¬Ü┴óū„ė├ÖCųŲĄ─╗»║Ž╬’Ż¼ą╬│╔ČÓ╬╗³cū„ė├Ż¼Ųõųą╚╬║╬ę╗ĘN╗»║Ž╬’ī”└źŽxĄ─▀xō±ē║┴”Č╝Ą═ė┌┐╣ąį░l(f©Ī)š╣Ą─ę¬Ū¾Ż¼ę“Č°▓╗Ģ■ę²░l(f©Ī)┐╣ąįĄ─«a(ch©Żn)╔·ĪŻÜóŽxä®╗ņė├Ę└ų╬║”ŽxĢrŻ¼ę╗ĘNĮMĘų▓╗─▄Üó╦└Ą─éĆ¾w┐╔─▄▒╗┴Ēę╗ĮMĘųÜó╦└Ż¼ę“×ķī”╦∙ėąÜóŽxä®Č╝Š▀ėą┐╣ąįĄ─éĆ¾wÄū║§▓╗┤µį┌ĪŻÜóŽxä®╗ņė├ū„×ķ┐╦Ę■╗“čėŠÅ┐╣ąį░l(f©Ī)š╣Ą─┤ļ╩®ąĶūóęŌęįŽ┬Äū³cŻ║ó┘╗ņė├Ą─Ė„ĮMĘųĄ─ū„ė├ÖCųŲæ¬(y©®ng)▒╦┤╦▓╗═¼Ż¼╚ń╣¹ū„ė├ÖCųŲŽÓ═¼Ż¼ät║”Žx║▄┐╔─▄ī”╗ņä®ųąĄ─Ė„ĮMĘų«a(ch©Żn)╔·Į╗╗ź┐╣ąįŻ¼Ę┤Č°┤┘▀M┴╦┐╣ąį░l(f©Ī)š╣Ż╗ó┌║”Žxī”╗ņė├Ą─├┐ę╗ĮMĘųĄ─┐╣ąįÖCųŲæ¬(y©®ng)įō▓╗═¼Ż¼╝┤║”Žxī”╗ņė├Ė„ĮMĘųĄ─┐╣ąįÖCųŲ▓╗═¼Ż╗ó█╗ņė├Ą─Ė„ĮMĘųĄ─Üłą¦Ų┌æ¬(y©®ng)╗∙▒ŠŽÓĄ╚Ż¼╚ń╣¹╗ņė├Ą─ĮMĘųų«ę╗Üłą¦Ų┌├„’@ķLė┌Ųõ╦¹ĮMĘųŻ¼īóĢ■įņ│╔▀xō±ē║┴”▓╗ŲĮ║ŌŻ¼╩¦╚ź╗ņė├Ą─ęŌ┴xĪŻ ÜóŽxä®▌åė├Ą─ų„ę¬ę└ō■(j©┤)╩Ū║”Žx┐╣╦Äąį╩Ūį┌ÜóŽxä®┤µį┌Śl╝■Ž┬Ī░╦▓ķg▀M╗»Ī▒Ą─ĮY(ji©”)╣¹Ż¼Ą½┐╣ąįéĆ¾wį┌ī”ÜóŽx䮫a(ch©Żn)╔·┐╣ąįĄ─═¼Ģr═∙═∙Ģ■«a(ch©Żn)╔·▀m║ŽČ╚┤·ārŻ¼╚ń░l(f©Ī)ė²Ų┌čėķLĪó╔·ų│┴”Ž┬ĮĄĪóéĆ¾wūāąĪĪó¾wųž£p▌pĄ╚Ż╗«ö(d©Īng)«a(ch©Żn)╔·┐╣ąįĄ─Ī░╦▓ķg▀M╗»Ī▒Śl╝■Ż©ÜóŽx䮥─▀xō±ē║┴”Ż®Ž¹╩¦ĢrŻ¼┐╣ąįéĆ¾wę“×ķ╔·╬’īW(xu©”)╔ŽĄ─┴ėä▌Č°ųØu▒╗╠į╠ŁŻ¼į┌ĘN╚║ųąĄ─Ņl┬╩Ž┬ĮĄŻ¼ūŅĮK┐╣ąįĘN╚║ī”╦Ä䮥─├¶ĖąČ╚Ģ■╗ųÅ═(f©┤)ĪŻ▀MąąÜóŽxä®▌åė├Ģrę¬ūóęŌŻ║ūŅ║├▀xō±Š▀ėąžō(f©┤)Į╗╗ź┐╣ąįĄ─╦Ä䮯¼ęįą╬│╔Ę┤▀xō±ū„ė├Ż¼ėąą¦čėŠÅ╗“ūĶų╣║”Žx┐╣╦ÄąįĄ─░l(f©Ī)╔·Ż╗▀xė├ū„ė├ÖCųŲ▓╗═¼Ą─╦Ää®▀Mąą▌åė├Ż¼▒▄├Ōą╬│╔Į╗╗ź┐╣ąįŻ╗įO(sh©©)ų├║Ž└ĒĄ─▌åė├ķgĖ¶Ų┌Ż¼▀@╩ŪÜóŽxä®▌åė├─▄ʱ│╔╣”Ą─ĻP(gu©Īn)µIŻ¼▌åė├ķgĖ¶Ų┌ę╗Č©ę¬ūŃęį╩╣└źŽxĘN╚║ī”╔Žę╗┤╬╦∙╩╣ė├╦Ä䮥─┐╣ąįĄ├ĄĮ╗ųÅ═(f©┤)Ż╗ÜóŽx䮥─▌åė├═¼śėę▓æ¬(y©®ng)įōį┌╦Ää®ķ_╩╝æ¬(y©®ng)ė├Ą─įńŲ┌Š═īŹ╩®ĪŻ└²╚ńŻ¼į┌├▐č┴╠’ķgĘ└ų╬▀^│╠ųą┐╔ęį▌åōQ╩╣ė├ī”├▐č┴Ę└┐žą¦╣¹▌^║├Ż¼Ūę┼cą┬¤¤ēAŅÉÜóŽx䮤oĮ╗╗ź┐╣ąįĄ─ą┬ą═ÜóŽxä®ļp▒¹Łh(hu©ón)Žx§źŻ¼ęįčėŠÅ├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį░l(f©Ī)š╣ĪŻ 4.3 ║Ž└Ē╩╣ė├į÷ą¦ä® į÷ą¦ä®╩Ūę╗ŅÉ▒Š╔Ēī”║”Žxø]ėą╔·╬’╗Ņąį╗“╗Ņąį▌^Ą═Ą─╗»║Ž╬’Ż¼Ą½Ųõ┼cÜóŽxä®┬ō(li©ón)║Ž╩╣ė├║¾Ģ■╩╣ÜóŽx䮥─ČŠ┴”’@ų°į÷ÅŖĪŻį÷ą¦ä®┐╔ęį═©▀^ęųųŲŽÓĻP(gu©Īn)ĮŌČŠ┤·ųx├ĖĄ─╗ŅąįŻ¼Ž¹│²ę“┤·ųx┐╣ąįÖCųŲ«a(ch©Żn)╔·Ą─┐╣ąįŻ¼Å─Č°▀_(d©ó)ĄĮ╠ßĖ▀╦Äą¦Ż¼£p╔┘ė├╦Ä┴┐Ż¼čėŠÅ┐╣ąį░l(f©Ī)š╣Ą──┐Ą─ĪŻ┤·ųx┐╣ąį╩Ū├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮫a(ch©Żn)╔·┐╣ąįĄ─ųžę¬įŁę“Ż¼ę“┤╦║Ž└Ē╩╣ė├į÷ą¦ä®─▄ē“į┌ę╗Č©│╠Č╚╔ŽčėŠÅ├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį░l(f©Ī)š╣ĪŻ└²╚ńŻ¼╣∙╠ņ°PĄ╚░l(f©Ī)¼F(xi©żn)Ż¼į÷ą¦├诩piperonyl butoxideŻ¼PBOŻ®║═╚²▒Į╗∙┴ū╦ߧźŻ©triphenyl phosphateŻ¼TPPŻ®─▄ē“’@ų°╠ßĖ▀├▐č┴ī”▀┴Žx▀°║═ÓżŽxļ▀Ą─├¶ĖąąįŻ╗ChenĄ╚蹊┐▒Ē├„Ż¼PBO║═TPPā╔ĘNį÷ą¦ä®─▄ē“’@ų°╠ßĖ▀├▐č┴ī”▀╗Žx░ĘĄ─ČŠ┴”ĪŻę“┤╦Ż¼▒M╣▄PBO║═TPP┐╔─▄▓╗─▄ų▒Įėū„×ķį÷ą¦ä®╩╣ė├Ż¼Ą½╩Ū▀@ą®čąŠ┐ĮY(ji©”)╣¹▒Ē├„═©▀^║Ž└Ē╩╣ė├į÷ą¦ä®┐╔ęį▀_(d©ó)ĄĮ╠ßĖ▀ą┬¤¤ēAŅÉÜóŽxä®ī”├▐č┴Ą─Ę└ų╬ą¦╣¹Ż¼£p╔┘ė├╦Ä┤╬öĄ(sh©┤)Ż¼ĮĄĄ═▀xō±ē║┴”Ą──┐Ą─ĪŻ 4.4 │õĘų└¹ė├Ė„ĘNĘ└ų╬┤ļ╩®īŹ¼F(xi©żn)ŠC║ŽĘ└ų╬ ├▐č┴Š▀ėąĘ▒ų│╦┘Č╚┐ņĪóĘN╚║öĄ(sh©┤)┴┐┤¾Īó▀mæ¬(y©®ng)ąįÅŖĄ─╠ž³cŻ¼─▄┐ņ╦┘▀mæ¬(y©®ng)Ę└ų╬╦Ää®▓ó«a(ch©Żn)╔·┐╣ąįŻ¼į┌╠’ķg├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąį░l(f©Ī)š╣╩«ĘųčĖ╦┘ĪŻę“┤╦Ż¼ę¬Å─Ė∙▒Š╔ŽĮŌøQ├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįå¢Ņ}Ż¼ąĶę¬┤¾┴”ķ_░l(f©Ī)║═└¹ė├▐r(n©«ng)śI(y©©)Ę└ų╬Īó╔·╬’Ę└ų╬Ą╚Łh(hu©ón)Š│ėč║├ą═┐žųŲ▓▀┬įŻ¼Ė∙ō■(j©┤)ŠC║ŽĘ└ų╬Ą─└Ē─Ņī”Ųõ▀MąąŠC║ŽĘ└ų╬ĪŻ▒M┴┐ĮĄĄ═├▐č┴ĘN╚║öĄ(sh©┤)┴┐Ż¼£p╔┘├▐č┴▒®░l(f©Ī)│╔×─(z©Īi)Ą─┐╔─▄Ż¼ĮĄĄ═ī”╗»īW(xu©”)▐r(n©«ng)╦ÄĄ─ę└┘ćąįĪŻ 05Īóš╣═¹ ─┐Ū░Į^┤¾ČÓöĄ(sh©┤)蹊┐āH─▄ūC├„P450ĪóUGTĄ╚┤·ųx├ĖĮŌČŠ┤·ųxį÷ÅŖ┼c├▐č┴ī”ą┬¤¤ēAŅÉÜóŽxä®┐╣ąįĄ─ŽÓĻP(gu©Īn)ąįŻ¼ų▒ĮėĄ─┤·ųxįć“×ūCō■(j©┤)▀Ć▒╚▌^╔┘Ż¼āHéĆäe╗∙ę“Ą─▒Ē▀_(d©ó)«a(ch©Żn)╬’═Ļ│╔┴╦┤·ųxįć“ׯ¼ę“┤╦▀ĆąĶ╝ėÅŖī”Ųõų▒Įė┤·ųxū„ė├Ą─蹊┐ĪŻ┤╦═ŌŻ¼─┐Ū░ļmėąP450Ą╚┐╣ąįŽÓĻP(gu©Īn)╗∙ę“▀^▒Ē▀_(d©ó)Ęųūėš{(di©żo)┐žÖCųŲĄ─ł¾Ą└Ż¼Ą½├▐č┴ī”ą┬¤¤ēAŅÉÜóŽxä®┐╣ąįą╬│╔Ą─ŽÓĻP(gu©Īn)╗∙ę“▀^▒Ē▀_(d©ó)Ęųūėš{(di©żo)┐žÖCųŲ▀Ć▌^╔┘Ż¼╚įąĶ╝ėÅŖŽÓĻP(gu©Īn)蹊┐ĪŻ ▒M╣▄Į³─ĻüĒėąļp▒¹Łh(hu©ón)Žx§źĪóĘ·ÓżŽx░ĘļµĄ╚ą┬ą═ÜóŽxä®æ¬(y©®ng)ė├ė┌├▐č┴Ą─╠’ķgĘ└ų╬Ż¼Ą½ą┬¤¤ēAŅÉÜóŽxä®╚į╩Ū╬ęć°─┐Ū░├▐č┴Ę└ų╬ųąæ¬(y©®ng)ė├ūŅČÓĄ─ę╗ŅÉÜóŽxä®ĪŻę“┤╦Ż¼├▐č┴ī”▀┴Žx▀°ĪóÓżŽxļ▀Ą╚ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįå¢Ņ}╚į▓╗╚▌║÷ęĢŻ¼ąĶę¬└^└m(x©┤)ī”Ųõ┐╣╦ÄąįÖC└Ē▀Mąą╔Ņ╚ļ蹊┐ĪŻį┌├▐č┴╠’ķgĘ└ų╬║═┐╣╦Äąįų╬└Ē▀^│╠ųąŻ¼ę¬│ų└m(x©┤)▒O(ji©Īn)£y├▐č┴╠’ķgĘN╚║Ą─┐╣╦Äąį¼F(xi©żn)ĀŅŻ¼▓óĖ∙ō■(j©┤)┐╣╦Äąį▒O(ji©Īn)£yĮY(ji©”)╣¹Ż¼ųŲČ©║Ž└ĒĄ─┐╣╦Äąįų╬└Ē▓▀┬įŻ¼┐ŲīW(xu©”)ųĖī¦(d©Żo)╠’ķgė├╦ÄŻ¼╠ßĖ▀╠’ķgĘ└ų╬ą¦╣¹ĪŻ═¼ĢrŻ¼ę¬Ė∙ō■(j©┤)├▐č┴ī”ą┬¤¤ēAŅÉÜóŽx䮥─┐╣ąįÖCųŲŻ¼į┌│õĘų└¹ė├¼F(xi©żn)ėąĘ└ų╬╦Ä䮥─╗∙ĄA(ch©│)╔ŽŻ¼ĮY(ji©”)║Ž▐r(n©«ng)śI(y©©)Ę└ų╬Īó╬’└ĒĘ└ų╬║═╔·╬’Ę└ų╬Ą╚ŠC║ŽĘ└ų╬ĘĮĘ©Ż¼▒M┴┐ĮĄĄ═ą┬¤¤ēAŅÉÜóŽxä®ī”├▐č┴Ą─▀xō±ē║Ż¼ęįčėŠÅ┐╣ąį░l(f©Ī)š╣Ż¼▀_(d©ó)ĄĮī”├▐č┴▀Mąą│ų└m(x©┤)Ę└┐žĄ──┐Ą─ĪŻ |
 |
 |
| ŽÓĻP(gu©Īn)ķåūx |
| ─├ö│ĘĆ(w©¦n)│╔ųąć°õN┴┐ūŅ┤¾Ą─ÜóŠ·ä®Ż¼┐╔Ę└ų╬ČÓĘNū„╬’ų„꬚µŠ·▓Ī║” |
| ▓╗┤“╦ÄĄ─▓▌▌«Ż¼─Ńą┼åß? |
| 2012─Ļ╗»īW(xu©”)▐r(n©«ng)╦ÄÜóŽxä®ŅA(y©┤)ėŗąĶŪ¾ |
 |
| ╩ūĒō | ┐ņėŹ | «a(ch©Żn)ŲĘÄņ | ÅS╝ę?gu©®)?/a> | łDūV | ĘNų▓╝╝Ū╔ |